Jest rzeczą ciekawą, że ta energetyczna, hamująca rola półkul mózgowych jest nie tylko najpierwotniejszą ich czynnością, ale także najtrwalszą i najbardziej powszechną, ponieważ stwierdzamy ją za równo u ryb kostno-szkieletowych jak w ogóle u wszystkich kręgowców aż do najwyżej stojących ssaków i człowieka. Mamy tu do czynienia z jakimś elementarnym, czysto fizjologicznym mechanizmem, z jedyną czynnością półkul mózgowych, która jest powszechna dla wszelkich postaci płaszcza mózgowego bez. względu na stopień jego rozwoju i zróżnicowania. Ta elementarność i ta powszechność czynności półkul, hamującej wszelkie dynamizmy nerwowe, leżące poniżej płaszcza mózgowego, zdaje się przemawiać za tym przyczynowym związkiem pomiędzy parciem ku przodowi dynamizmów podkorowych a powstawaniem płaszcza mózgowego, o którym przed chwilą mówiliśmy jako o biologicznej potrzebie stworzenia nowego akumulatora energii nerwowej. To właśnie parcie jest chronologicznie najwcześniejszym przejawem wędrówki czynności ku przodowi, albo ściślej mówiąc, wędrówka niezróżnicowanej energii nerwowej ku przodowi jest zjawiskiem ewolucyjnie wcześniejszym niż wędrówka energii nerwowej zróżnicowanej.
Rola płaszcza mózgowego jako akumulatora energii i globalnego hamulca występuje wyraźnie już u tych niższych kręgowców, których podkorowe nastawienia instynktowe bynajmniej jeszcze nie straciły swych bocznych członów. Jeden z nich umożliwia rozpoznanie bodźca biologicznie ważnego, drugi zaś umożliwia reakcję ruchową w znaczeniu klisis lub ekklisis, przyciągania i odtrącania.
Ale i po powstaniu tego elementarnego płaszcza mózgowego wywierane nań parcie dynamizmów podkorowych trwa dalej i powoduje dalszą jego ewolucję, która anatomicznie wyraża się w coraz dalej posuwającym się różnicowaniu pól cytoarchitektonicznych i tworzeniu się zróżnicowanego, swoistego przedstawicielstwa w korze mózgowej dróg zmysłowych, czucia głębokiego i dróg ruchowych. Jeżeli zgodzimy się z K. Brodmannem na jego twierdzenie, że »czynność tworzy sobie swoje narządy», i zapytamy, jaka to czynność, tworzy zróżnicowane, swoiste narządy zmysłów i ruchowości w korze mózgowej, to zdaje się, że odpowiedź na to rzeczywiście może być tylko jedna: dalszy ciąg dróg projekcyjnych w korze mózgowej powstaje pod wpływem działania na korę dynamizmów talamicznych, które ze swej strony powstają z przekształcania energii podrażnień właśnie wszystkich dróg dośrodkowych poprzez jednostki rdzeniowo-segmentarne; za takim właśnie mechanizmem powstawania pierwszych zróżnicowanych narządów korowych przemawia przede wszystkim proces tak zwanej wędrówki czynności ku przodowi, polegającej na przejmowaniu przez korę mózgową czynności podkorowych, w biologicznym celu ich udoskonalenia. Gdy się mówi o wędrówce czynności ku przodowi, ma się zwykle na myśli czynności rozpoznawania za pośrednictwem zmysłów i czynności ruchowe, czyli czynności uważane zazwyczaj za czynności układu projekcyjnego, wraz z jego korowym przedstawicielstwem. Ale jest rzeczą wiadomą, że płazy, gady, ryby i ptaki nie mają jeszcze dróg piramidowych, że bezmózgi np. gołąb jeszcze widzi, omija przeszkody, chociaż wykazuje utratę pamięci i nie rozpoznaje widzianych przedmiotów, gołębica nie zważa na wabienie samca i nie troszczy się o swe pisklęta, które za nią gonią, jest w ogóle obojętna i senna (A. Beck ). Kora mózgowa ptaków jest na podrażnienia laboratoryjne jeszcze niepobudliwa. Zanik widzenia po wycięciu półkul mózgowych lub płatów potylicznych występuje dopiero u ssaków, np. u opossum, chociaż Lashley podkreśla, że jeszcze gryzoń pozbawiony płatów potylicznych nie tylko odróżnia światło od ciemności, ale zdolny jest do poznania przedmiotu i jego odległości, co mu pozwala omijać przeszkody, podczas gdy zupełne przecięcie włókien wzrokowych wywołuje skutki gorsze, zupełnie uniemożliwiające zwierzęciu orientowanie się za pośrednictwem wzroku.
Jak widzimy, całkowite czy też prawie całkowite przeniesienie czynności zmysłowych i ruchowych z mechanizmów podkorowych na korę mózgową występuje dopiero na bardzo wysokim szczeblu rozwojowym, mianowicie u ssaków, natomiast u kręgowców niższych przedstawicielstwo układu zwierzęcego w płaszczu mózgowym zdaje się jeszcze w ogóle nie istnieć. A jeśli tak jest, to zachodzi pytanie, z jakiej tkanki nerwowej zbudowany jest ten prymitywny płaszcz mózgowy kręgowców niższych, który jest akumulatorem energii nerwowej niezróżnicowanej i hamulcem nadmiernej ruchliwości podkorowej, jak to wykazuje zmniejszona ostrożność zwierząt, pozbawionych półkul mózgowych. Zdaje się, że odpowiedź nie może tu budzić nadmiernych wątpliwości, bo jeżeli kora mózgowa ryby kostno-szkieletowej nie posiada w ogóle żadnego układu nerwowego zwierzęcego, to oczywiście może się na nią składać tylko układ nerwowy drugi, mianowicie wegetatywny. Zresztą za tym przypuszczeniem przemawia także jego charakter akumulatora, gromadzącego energię nerwową niezróżnicowaną, pochodzącą z podrażnień dośrodkowych, przerobionych w ośrodkach rdzenia kręgowego i we wzgórzach wzrokowych, ale wykazujących już na obwodzie zróżnicowanie swoiste bardzo wyraźne. Należy również podkreślić, że wegetatywny charakter płaszczów mózgowych rozwojowo najniższych zgadza się w zupełności z ogólnym planem budowy i czynności układu nerwowego ośrodkowego, mianowicie z rolą środkowego ogniwa w wielkim łuku, pomiędzy talamicznymi zespołami podrażnień dośrodkowych a narządami ruchowymi, czyli z rolą czynnika kierującego czynnościami układu nerwowego zwierzęcego. Zresztą wegetatywny charakter płaszcza mózgowego kręgowców niższych podkreślany jest już dość powszechnie. Ze swej strony możemy jeszcze zaznaczyć analogię płaszcza mózgowego, jako akumulatora i transformatora dopływającej energii nerwowej, z tym ośrodkowym, wegetatywnym ogniwem łuku odruchowego rdzeniowego, w którym neurofizjologia stwierdza również zjawisko kumulowania i przekształcania energii podrażnień dopływających, znane pod nazwą »sumowania się podrażnień». W obu tych przypadkach mamy do czynienia ze zjawiskiem o charakterze bardzo elementarnym, ponieważ w obu przypadkach zostaje kumulowana energia nerwowa nieswoista, niezróżnicowana, bez względu na ewentualną swoistość podrażnień dopływających. W tym oświetleniu filogenetyczny rozwój korowego przedstawicielstwa układu zwierzęcego, projekcyjnego, odbywa się już nie na tle korowego układu wegetatywnego, ale wręcz w samej jego rozlanej masie, tworząc w korze mózgowej nierozerwalny splot obu układów nerwowych, wegetatywnego i zwierzęcego. Stąd może również bardziej zrozumiała jest cecha nieswoistości i powszechności wegetatywnego akumulatora korowego. Oczywiście nie jesteśmy i zapewne nigdy nie będziemy w możności powiedzieć, czy z dynamiczną nieswoistą czynnością tego wegetatywnego akumulatora wiąże się już jakieś »fizjologiczne uczucie» przyjemności i przykrości, używając wyrażenia Monako w a i Mourgue’a. Ale sądząc ze wszystkiego, co było mówione w tej pracy o stosunku życia uczuciowego do układu wegetatywnego, taka hipoteza wydaje się raczej prawdopodobna, zwłaszcza że ten korowy układ wegetatywny musimy uważać rozwojowo przede wszystkim jak gdyby za dalszy ciąg talamicznego ośrodka afektywności protopatycznej i podkorowych ośrodków potrzeb fizjologicznych.
Różnicowanie jednolitej kory mózgowej przez tworzenie w niej nowych narządów poznania i ruchu odbywa się zatem według naszej »hipotezy roboczej», przez aktywność własną układu wegetatywnego, psychologicznie odpowiadającą tendencjom ustroju do ponownego zdobycia tych bocznych członów (gnostycznego i kinestetycznego) każdego pierwotnego zespołu mnemicznego instynktu, które ulegają zanikowi zwłaszcza w filogenezie ssaków.
Powstawanie nowych tendencji, nowych nastawień poznawczo-uczuciowych, dających się stwierdzić przez nowe reakcje (odruchy warunkowe), jest równoznaczne z powstawaniem nowego typu hamulców korowych i aktywności korowych, nowego dlatego, że ma on charakter wybiórczy, swoisty, ściśle ograniczony — podobnie jak uczucie każdej potrzeby fizjologicznej — a więc przeciwstawny elementarnemu, ogólnemu hamowaniu wszelkich czynności mechanizmów niżej położonych, przez korę i jej energię niezróżnicowaną. Musimy powiedzieć, że ten nowy typ nastawień korowych, jak już wiemy, posiadających również charakter energetyczno-mnemiczny, jest nowym typem energii nerwowej, mianowicie zróżnicowanej, ale równocześnie i zsyntetyzowanej, ponieważ powstaje ze zlania się w jedną całość czynnościową (zespół poznawczo-uczuciowy) obu jego elementarnych składników mnemicznych, wegetatywnego i zwierzęcego. Ani składnik uczuciowy nie może się rozwijać bez równoczesnej jego coraz dalszej intelektualizacji, ani składnik gnostyczny nie może ulegać ewolucji bez równoczesnej pracy jakiejś tendencji uczuciowej i uwagi; dalszy rozwój jest rozwojem tych zespołów, coraz bardziej oderwanych i odległych od popędów instynktowych i od spostrzegania konkretnego, i doprowadzających w końcu do powstania trzeciego, najwyższego typu hamulców korowych i aktywności korowej, mianowicie czołowo sprzężonego charakteru. Pomiędzy reaktywnymi czynnościami natury odruchowo-warunkowej a wewnątrzkorowymi czynnościami narządu czołowego istnieje pośredni okres czynności wewnątrzkorowych pozaczołowych (dynamizmy prelogiczne). Czynności intrapsychiczne (wewnątrzkorowe) obu wielkich poziomów ewolucyjnych zdolne są już nie tylko do aktywności własnej hamującej, ale co więcej, także do aktywności własnej, mającej charakter inicjatywy samorzutnej, będącej możliwie najwyższą postacią dynamizmu psychofizjologicznego. Trzy typy hamowania, o których tu mówiliśmy, są czynności ą dwóch wielkich narządów korowych, ponieważ okres reaktywności warunkowej i aktywności wewnątrz korowej elementarnej musielibyśmy zaliczyć do dynamizmów narządu pozaczołowego. Również możemy wykryć tylko dwa źródła, z których bezpośrednio wypływają dynamizmy korowe, jednym jest fizjologiczna konstelacja na obwodzie (zwłaszcza stan napełnienia żołądka i pęcherzyków nasiennych), który jest głównym motorem zachowania się zwierząt, drugim zaś jest konstelacja intrapsychiczna, która u człowieka dochodzi do wysokiego rozwoju i do inicjatywy.
Jeżeli wróciliśmy tutaj raz jeszcze do zagadnienia ewolucji czynności korowo-psychicznych, to stało się to dlatego, aby uprzytomnić sobie z całą wyrazistością, że to, co nazywamy synergią dynamizmów nerwowych (lub psychicznych), przedstawia się w sposób bardzo różny, zależny od okresu rozwoju tych dynamizmów.
Synergia ta jest na najniższych szczeblach rozwoju kręgowców zawsze idealną harmonią pomiędzy obwodem i ośrodkami nerwowymi. Harmonia ta nie może nigdy ulec zakłóceniu dlatego, że obwód i ośrodki stanowią zupełnie jednolity układ energetyczny, ponieważ podkorowe ośrodki instynktu mogą być czynne tylko pod wpływem działania bodźców na obwodzie, albo nawet, jak w przypadkach potrzeb fizjologicznych, są wręcz naładowywane energią powtarzających się podrażnień dośrodkowych ustrojowych. Układ nerwowy energetyczny staje się z natury rzeczy dużo bardziej pojemny, gdy ewolucja dołącza do pierwotnego, podkorowego układu najpierw płaszcz mózgowy wegetatywny, a potem jeszcze swoisty narząd pozaczołowy; ale na podstawie doświadczeń z odruchowością warunkową u zwierząt wyższych i u dziecka możemy powiedzieć, że i ten nowy, bardziej pojemny układ energetyczny wciąż pozostaje jeszcze jednolity, gdyż synergia jego dynamizmów bynajmniej nie psuje dawniejszej doskonałej harmonii. Musimy upatrywać w trwaniu tej harmonii dowód, że nowy, pozaczołowy dynamizm, który określaliśmy jako nowe, udoskonalone kierownictwo, umożliwiające lepsze dostosowywanie się organizmu do zmiennych warunków świata zewnętrznego, w gruncie rzeczy jest zależny od zupełnie tych samych sił, które wchodziły w grę w mechanizmach podkorowo-instynktowych, a więc od konstelacji ustrojowej na obwodzie i od nastawień mnemiczno-gatunkowych instynktu. Rzutowanie talamicznych zespołów na korę pozaczołową umożliwia intelektualizowanie, osłabianie, wzmacnianie, przekształcanie poszczególnych dążności instynktowych, ale mimo wszystkie te zmiany mamy tu w gruncie rzeczy do czynienia wciąż z tymi samymi podstawowymi motorami dynamicznymi, które znajdujemy w instynkcie. Zwierzęta i dziecko, wykazujące już nowe odruchy warunkowe, pozostają jednak na ogół wciąż jeszcze istotami instynktowymi w znaczeniu bezpośredniości reakcji i rodzaju samych dążności, wśród których uczucia iradiowane nie zyskują jeszcze żadnej samodzielności. Krótkie chwile wahania u dziecka 8-miesięcznego Watsona, możliwe tylko tam, gdzie dążności uczuciowe zostały już o tyle intelektualizowane, że umożliwiają kojarzenie zespołów poznawczo-uczuciowych i wzajemne ich wpływanie na siebie — dowodzą niewątpliwie, że zaburzenia harmonii i jedno-znacznej postawy synergetycznej mogą występować przejściowo już w drugim półroczu życia dziecka jako zjawisko zupełnie normalne — ale w tym wieku wahania takie są zawsze współzawodnictwem dwóch sił instynktowych, rzutowanych na korę. Co więcej, nie tylko odruchowość warunkowa, ale nawet dynamizmy prelogiczne, stojące już ewolucyjnie wyżej od odruchowości warunkowej, jako z niej wyrastające i posiadające charakter automatyzmów intrapsychicznych, bynajmniej nie psują tej pierwotnej jednolitości energetycznej ustroju dziecka, a w pewnych warunkach mogą nierzadko nie wykazywać nawet takich przejściowych wątpliwości i wahań, jakie powstają niekiedy w sytuacjach realnych, pod wpływem bodźców konkretnych.
Wracamy więc znów do stwierdzenia tego, że dualizm energetyczny, psujący jednolitość i harmonię pierwotną, psychologicznie i neurologicznie jako zjawisko trwałe i ciągłe, przejawiające się neurologicznie w istnieniu narządu korowego pozaczołowego i czołowego, psychologicznie zaś w równoległym istnieniu w świadomości dwóch jej prądów — myślenia konkretnego (lub prelogicznego) i czołowo-logicznego — występuje dopiero wówczas, gdy rozwinie się dynamizm czołowy, czyli psychologicznie gdy rozwiną się te najwyższe warstwy i dynamizmy psychiczne, które wyróżniają człowieka od zwierzęcia. Fakt, że najwyższe warstwy psychiczne rozwijają się tylko tam, gdzie istnieje narząd mowy, wskazuje na przy-czynowy związek pomiędzy tymi dwiema kategoriami zjawisk. Związek ten jest dwojakiego rodzaju. Psychologiczny związek jest okrężny: wzrastająca inteligencja dąży do porozumiewania się z innymi ludźmi, i powstały stąd narząd mowy staje się znakomitym środkiem porozumiewania się z innymi ludźmi, zdobywania dla siebie ich wiedzy i doświadczenia, wywierania wpływu moralnego itd., a więc rozwoju inteligencji i życia uczuciowego, powstawania nowych nastawień intencjonalnych, uspołecznionych i myślenia przyczynowo-logicznego. Anatomiczny narząd mówienia zdaje się pozostawać jeszcze w zupełnie innym związku przyczynowym z tworzeniem się narządu czołowego, mianowicie w związku, zależnym od stosunków topograficznych i energetycznych.
Wzmiankowaliśmy już o tym, że globalne dynamizmy nerwowe, jakościowo różne, zdają się istnieć tylko tam, gdzie są od siebie wyraźnie przestrzennie oddzielone, jak np. narząd pozaczołowy od pod- korowego przez całą długość dróg coronae radiatae, albo narząd czołowy od pozaczołowego przez okolicę ruchową (wraz z narządem mówienia). Zdaje się, że mamy tu do czynienia z jakąś regułą ogólniejszą, jeżeli można sądzić z tego, że i w obrębie tego samego wielkiego narządu korowego znajdujemy stosunki analogiczne pomiędzy poszczególnymi »ośrodkami korowymi». Mamy tu na myśli to, co Potzl i Hoff nazywają »czynnością poprzeczną», która zdaje się być mechanizmem koniecznym tam, gdzie dwa odrębne dynamizmy, lokalne lub globalne, nie są od siebie odseparowane przestrzennie w sposób wystarczający, tak jak narząd pozaczołowy od podkorowego.
Dynamizmy czołowo-logiczne tak się rozwijają, jak widzieliśmy, z dynamizmów popędowo-prelogicznych, jak odruchowość warunkowa z instynktu. Anatomicznym warunkiem tego rozwoju jest istnienie i prawidłowa czynność narządu czołowego i narządu mowy. Wiadomo jest, jak zwłaszcza w literaturze anglosaskiej podkreślana jest równoległość w stopniowaniu niedorozwojów oligofrenicznych i niedorozwojów mowy. Narząd ruchu i mowy zdaje się w ogólnym planie działania kory mózgowej odgrywać rolę takiej »funkcji poprzecznej» pomiędzy narządem pozaczołowym a czołowym, jaką odgrywa w obrębie pozaczołowym np. zwój międzyciemieniowy, dzielący i wiążący sferę dotyku ze sferą wzrokową.
Mówiąc o swoistych czynnościach kory mózgowej, nie mamy prawa przeciwstawiać całości płaszcza mózgowego czynnościom instynktowym, ponieważ stosunek do nich narządu pozaczołowego i narządu czołowego jest zupełnie inny, świadczący właśnie o odrębności ich dynamizmów. Jest coś nie tylko naiwnego, ale i szkodliwego w dążności do poznania czynności anatomicznego narządu świadomości bez wzmiankowania o czynnościach psychicznych, albo w określaniu ich jakimiś nowymi, nic nie mówiącymi terminami fizjologicznymi, ponieważ stanowisko takie utrudnia zrozumienie nie tylko zjawisk psychicznych, ale i fizjologii kory mózgowej. Możemy wprawdzie powiedzieć językiem wyłącznie fizjologicznym, że narząd pozaczołowy jest narządem odruchowości warunkowej, narząd zaś czołowy jest zwierzchnim kierownictwem narządu pozaczołowego, narządu podkorowego i narządu móżdżkowego; ale jest rzeczą jasną, że nie daje to żadnego właściwego wyobrażenia o tych narządach, które są w istocie swej i przede wszystkim narządami dwojakiego rodzaju: aktywności psychicznej, ewolucyjnie niższej i ewolucyjnie wyższej.
Z powyższego wynika, że synergia układów nerwowych noworodka jest synergią mechanizmów obwodowych z gatunkowymi mechanizmami podkorowymi. U dziecka jest synergią mechanizmów obwodowych, jeszcze żywo działających podkorowych nastawień instynktowych i coraz bardziej uczulających się i mnożących nastawień poza- czołowych, pobudzających i hamujących. Wreszcie u człowieka dojrzałego podkorowe czynności »czystego» instynktu spadają tak nisko, że prawie nie wschodzą w rachubę. Nie znaczy to bynajmniej, że popędy instynktowe przestają działać u człowieka dojrzałego, gdyż w rzeczywistości działają do końca życia, do późnej starości, ale pod postacią zespołów o nastawieniach pozaczołowych, uświadamianych, mniej lub więcej intelektualizowanych. W każdym razie w wieku dojrzałym znajdujemy już stale energetyczną dwoistość, przejawiającą się fizjologicznie w synergii dominanty pozaczołowej i dominanty czołowej, pozostających w stanie równowagi ruchomej, psychologicznie zaś w dwoistości równoległych prądów świadomości — życia ustrojowo- przedmiotowego lub myślenia prelogicznego i życia czołowo-logicznego.
W tej synergii mamy do czynienia wciąż z wzajemnym oddziaływaniem wszystkich tych dynamizmów, skąd wynika ich wpływ okrężny. A ponieważ układ nerwowy ośrodkowy możemy uważać za układ energetyczny w pewnej mierze zamknięty (podobnie jak ustrój żyjący w stosunku do świata zewnętrznego), czyli rozporządzający pewną ograniczoną ilością energii nerwowej, więc każda zmiana w którymkolwiek narządzie piętrowym musi z natury rzeczy wywołać odpowiednie zmiany w energetycznym stanie pięter innych.
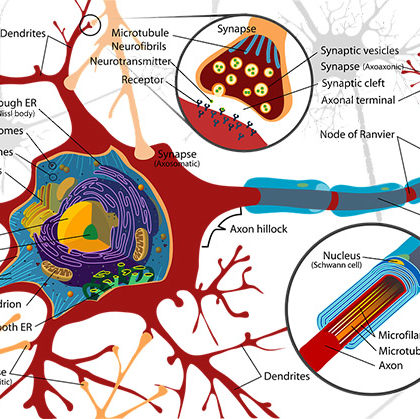
Zdjęcie pochodzi z portalu flickr.com. Autorem grafiki jest Rawpixel Ltd. Zdjęcie zostało wykorzystane na podstawie licencji CC BY 2.0.