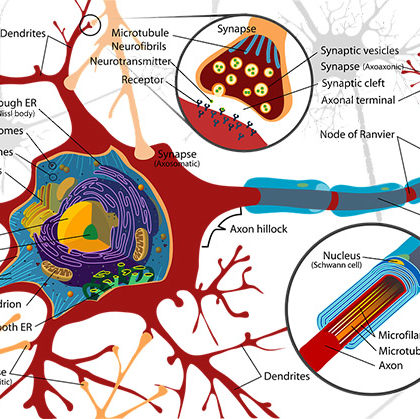
Z poprzednich artykułów wynika, że cały rozwój życia psychicznego w wąskim tego słowa znaczeniu, tj. bez instynktu, odbywa się od początku do końca wyłącznie w obrębie kory mózgowej. Natomiast wszelkie źródła jego dynamiki są oczywiście natury pozakorowej, ponieważ istnieje napęd ze strony układu gruczołów dokrewnych i napęd — niezróżnicowany i zróżnicowany — pochodzenia nerwowego. Z tego punktu widzenia można twierdzić nie bez pewnej słuszności, że czynności psychiczne »lokalizują się» w całym ustroju, — ale z jeszcze większą słusznością możemy twierdzić, że wszystko, co jest swoiste, co różni czynności psychiczne od »czysto» neurologicznych, tj. tylko cielesnych, powstaje jedynie i wyłącznie w płaszczu mózgowym, w dwóch jego wielkich narządach korowych, przekształcających energię dopływających dynamizmów talamicznych na swój własny, swoisty sposób, odpowiadający właściwościom budowy anatomiczno-fizjologicznej danego piętra. Każdemu takiemu tworzeniu nowych swoistych czynności nerwowych, czyli ich różnicowaniu towarzyszy zawsze proces scalania, czyli wcielania tych nowych nabytków czynnościowych do poprzedniej całości, która na nowym poziomie odpowiednio przekształca się; albo ściślej mówiąc, zróżnicowaniu ulega zawsze całość, która zatem — pomimo dokonanej przez zróżnicowanie zmiany — pozostaje całością i jednością czynnościową zawsze, we wszystkich stadiach rozwoju ustroju i żadna z tych jedności nie jest nigdy sumą poszczególnych swych składników. Powtarzamy raz jeszcze dlatego, że fakty doświadczalne bardzo ścisłe, na których oparty jest ten nowy pogląd psychofizjologiczny, są, naszym zdaniem, ogromnej doniosłości, ponieważ: 1) wbrew dawniejszym poglądom stwierdzają odrębną jakość dynamizmów poszczególnych pięter, zgodnie z tymi poglądami (Claparède, Piaget), które widzą w wielkich stadiach rozwoju psychiki różnice nie tylko ilościowe, ale i jakościowe, 2) przenoszą nas z ciasnej dziedziny neuronów, łuków odruchowych i przewodnictwa podrażnień w poszczególnych neuronach w nową dziedzinę globalnych dyna m izmów piętrowych coraz bardziej złożonych, w dziedzinę nieznaną nam dotąd bliżej, ale pomimo tego stwierdzoną i bliższą życia realnego niż »wygodna fikcja» odruchu, i 3) umożliwiają względnie lepsze fizjologiczne zrozumienie zasad ewolucyjnych i dysolucyjnych Hughlings Jacksona, wyprowadzonych z psychopatologii i neurologii. Jest koniecznością zrozumienie tego, że zasady Jack sona — wbrew jego własnej opinii — mogą być uzasadnione tylko w takim razie, jeżeli pomiędzy czynnościami i dynamizmami poszczególnych pięter nerwowych istnieją wyraźne różnice jakościowe, bez których ewolucja od odruchu do »woli» jest nie do pomyślenia, gdyż każdy odruch jest czynnością, która do niedawna była uzależniana wyłącznie od działania bodźca, czynności zaś dowolne uzależniane są zawsze od aktywności własnej, czyli od przeszłości, a nie od chwilowego bodźca. Siły świata zewnętrznego i dążności ustroju, wyrażające się w jego aktywności własnej, nie mogą być sprowadzone do różnicy tylko ilościowej. Ale rozbieżność całkowita istnieje chyba tylko w tych stosunkach np. u jamochłonów, których mieszana komórka odbiorczo-mięśniowa nie ma jeszcze nic wspólnego z siecią nerwową. Natomiast wszędzie tam, gdzie pomiędzy zróżnicowany już narząd nerwowy odbiorczy a- narząd nerwowy wykonawczy wciska się jako regulator i sternik wegetatywny układ nerwowy, »produkujący» aktywność własną, — tam znajdujemy już i pewne różnice ilościowe, mianowicie: wraz z ewolucją czynności i) zmniejsza się znaczenie nasilenia bodźca zewnętrznego, 2) zwiększa się znaczenie aktywności własnej, tj. dynamizmów pochodzenia mnemicznego, nawarstwiających się na siebie kolejno i tworzących tzw. »warstwy chronogenne», o których mówiliśmy w rozdziale o pamięci fizjologicznej. Tę nazwę »lokalizacji chronogennej» zastosował Monakow do nie mogących ulegać wątpliwości faktów natury psychologicznej i psychopatologicznej. Natomiast strona biologiczna tych zjawisk jest już dużo mniej wyraźna, a strona fizjologiczna jest zupełnie niejasna, trudno jest sobie nawet wyobrazić fizjologiczny mechanizm nawarstwiania się chronogennego. Względnie najłatwiej ten proces nawarstwiania się kolejnego możemy sobie wyobrazić, oczywiście, w przykładach engrafii najbardziej elementarnej, a więc w prostych zjawiskach sumowania się skutków podrażnień, np. w wegetatywnym ogniwie ośrodkowym łuku odruchowego rdzeniowego, albo w podkorowych ośrodkach, których naładowanie »produkuje» uczucie potrzeby fizjologicznej. Ale na poziomie kory mózgowej, tego szczytowego narządu engrafii zespołowej, trudno jest nawet wyobrazić sobie jakąś możliwość fizjologiczną kolejnego nawarstwiania się współczesnych zespołów wrażeń, oryginalnych i mnemicznych. Przypuszczenie Semona, że »engramy zostają przechowywane w chromatycznej istocie jąder komórkowych» zanadto przypomina »komórki psychiczne» i »komórki pamięciowe» ubiegłego wieku, słusznie dzisiaj odtrącane już powszechnie. Zdaje się, że dzisiaj możemy już nawet z punktu widzenia fizjologicznego powiedzieć, czemu żadna lokalizacja »komórkowa» — poza przykładami wspomnianych tu przed chwilą elementarnych zjawisk sumowania się skutków podrażnień — jest w korze mózgowej w ogóle niedopuszczalna. Mianowicie możliwość ta istniała tylko dla tych autorów, którzy czynności układu nerwowego i psychiki ujmowali z punktu widzenia teorii mechanistycznej, ale oczywiście możliwość przestaje istnieć dla tych autorów, którzy stoją na gruncie dynamistycznym, skoro widzieliśmy, że dynamizmy wielkich narządów piętrowych nie dają się sprowadzić do sumy podrażnień dopływających. Dynamizmy te bowiem stanowią odrębną, swoistą jedność czynnościową, która uzupełnia swe braki (wydatki) energetyczne z energii tych podrażnień dopływających, podobnie jak organizm żyjący czerpie swą energię ze świata zewnętrznego. W obu jednak tych przypadkach otrzymywana energia jest przekształcana na własną energię, zależną od budowy i fizyko-chemicznych właściwości tkanki odbierającej.