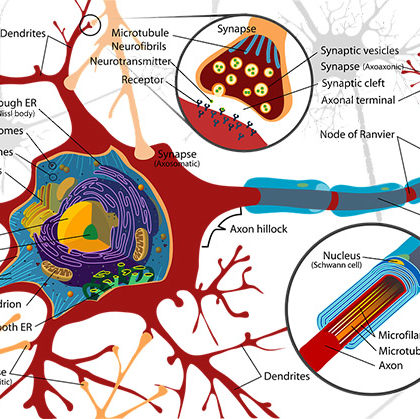
Powtarzająca się i nawet »spontaniczna» aktywność nie jest swoistą własnością tkanki nerwowej ośrodkowej, ponieważ jest spostrzegana bardzo łatwo na izolowanych neuronach obwodowych, po ich urazie, albo w innych warunkach, przeważnie anormalnych. Wprawdzie nie ma żadnego bezpośredniego dowodu na to, że pojedynczy neuron korowy jest zdolny do rytmicznych wyładowań bez okresowego ich uczynniania przez impulsy nerwowe dochodzące do synapsy tego neuronu, ale Jasper jest skłonny sądzić, że powolne spontaniczne fale w korze mózgowej mogą być potencjałami komórek podobnymi do tych, jakie Heinbecker spostrzegał na powierzchni ciała pojedynczych komórek w zwojach serca limuli, pomimo dużych różnic pomiędzy własnościami obu tych rodzajów komórek.
Ranson i Hinsey, Kubie, a także Lorent de Nó budują teorię łańcuchów neuronów ośrodkowych, tworzących zamknięte kręgi, wzdłuż których mogą krążyć impulsy nerwowe nieskończenie, dopóki każdy neuron w łańcuchu nie wypocznie dostatecznie przez ten przeciąg czasu, w którym musi być ponownie pobudzony, aby przewodzić impulsy do następnego neuronu w łańcuchu. Dowód na rzecz tej teorii kręgów wewnątrzkorowych spoczywa głównie na histologicznej demonstracji istnienia łańcuchów neuronowych, które mogą działać w ten sposób. Barthley i Bishop (1933), a także Bishop (1936) spostrzegali, że część kory wzrokowej królika może być prawie otoczona przez nacięcia, które jednak nie powodują zaburzeń aktywności elektrycznej, dopóki zachowane jest zaopatrywanie w krew, ale przecięcie pewnych dróg, prawdopodobnie tych, przez które impulsy nerwowe dochodzą do pola wzrokowego, powoduje zanik wszelkiej działalności. Przecięcie radiatio optica powyżej wzgórza wzrokowego znosi powolny rytm korowy 3—7 drgań na sekundę, pozostawiając tylko rytm drobny o 30—80 drganiach na sekundę. Stąd wnioskują ci autorzy, że ten ostatni rytm jest utrzymywany wewnątrz komórek korowych, większy zaś powolny rytm zależy od impulsów, krążących w drogach korowo-wzgórzowych i wzgórzowo-korowych, mających ty]) kręgu »odbijającego», który został wykazany przez Lorenta de Nó w funkcjonowaniu ośrodków przedsionkowo-ocznych. Nie można jeszcze dzisiaj określić, w jakich rozmiarach autonomiczna aktywność komórek korowych zależy od tego, czy »odbijające» kręgi funkcjonalne są nienaruszone, w jakich zaś rozmiarach zależy od spontanicznie rytmicznych właściwości pojedynczego neuronu, niezależnych od okresowego aktywowania. Nawet gdyby rytmy korowe przedstawiały powtarzające się wyładowania, odnoszące się do nie- rytmicznego »stanu stałego», bądź na podstawie chemicznej, jak przypuszcza Ho ag land (1935), bądź też na podstawie elektrotonicznej, jak to przypuszcza Jasper (1936), to takie hipotezy są według Lorenta de Nó nadmiernie uproszczone i przedwczesne, dopóki nie dowiemy się więcej o charakterze przewodnictwa impulsów w drogach ośrodkowych i korowo-rdzeniowych, korowo-talamicznych, spoidłowych, wewnątrzkorowych itd.
Pomijając jednak na razie wszelkie teorie i hipotezy, musimy z powyższego wnioskować, że istnienie w korze mózgowej ciągłych samorzutnych, rytmicznych wyładowań elektrycznych należy uważać za fakt dawno stwierdzony. Z naszego punktu widzenia musimy tu podkreślić, że ciągłość rytmicznych oscylacji jest cechą pospolicie spotykaną w aktywności układu nerwowego wegetatywnego, w przeciwstawieniu do układu zwierzęcego, którego część dośrodkowa pozostaje w ciągłym spoczynku, dopóki nie działa jakaś podnieta zewnętrzna, i część odśrodkowa pozostaje znów w ciągłym spoczynku, dopóki nie działa na nią bodziec ośrodkowy. Również »samorzutność», »spontaniczność», niezależność istnienia tych rytmów od podniet zewnętrznych lub ustrojowych, zmusza nas do zaliczenia tych rytmów do kategorii tej »aktywności własnej» układu nerwowego, z którą w rozdziałach poprzednich spotykaliśmy się wciąż przy omawianiu układu wegetatywnego, ale nie zwierzęcego. Oczywiście nie mamy tu na myśli obwodowych części układu wegetatywnego, pozostających w znanej ścisłej zależności od układu gruczołów dokrewnych, ale mówimy tu o układach wegetatywnych ewolucyjnie wyższych, a więc o wegetatywnym przedstawicielstwie wyższych pięter układu nerwowego, pozostających w stosunkowo znacznie mniejszej zależności od wpływów hormonalnych niż części obwodowe.
Powtarzające się spontaniczne wyładowania elektryczne musiałyby doprowadzić, według wzmiankowanego już zdania Adria- n a, do chaosu, gdyby istniały w neuronach ruchowych lub mięśniach szkieletowych, ale ten wniosek niekoniecznie musi dotyczyć neuronów ośrodkowych. Można przypuszczać, że doprowadziłyby także do chaosu, gdyby istniały w drogach dośrodkowych, np. zmysłowych, które są przeznaczone z natury swej do przewodzenia właśnie podrażnień nie spontanicznych, ale wywoływanych przez odpowiednie bodźce, i które zresztą pozostają — analogicznie do neuronów ruchowych — w spoczynku dopóty, dopóki nie działa bodziec. W takich warunkach wydaje się wątpliwe, aby korowe przedstawicielstwo układu dośrodkowego zwierzęcego mogło wykazywać wyładowania elektryczne spontaniczne i rytmiczne, skoro i spontaniczność, i rytmiczność, i ciągłość funkcjonowania są cechami obcymi dla układu zwierzęcego. Natomiast układ wegetatywny obwodowy pozostaje wprawdzie pod ciągłym wpływem czynników hormonalnych, ale ze swej strony posiada wszystkie te trzy cechy w wysokim stopniu; mianowicie i rytmiczność, począwszy od akcji serca i oddychania, a kończąc na odżywianiu, na miesiączce, na zmienności stanów snu i czuwania, — i ciągłą spontaniczność, przejawiającą się w ciągłości wszelkich czynności kierowniczych układu wegetatywnego na wszystkich jego piętrach. Stąd wynika prawdopodobieństwo, że powtarzające się wyładowania spontaniczne, stwierdzone w korze mózgowej, istnieją raczej w jej przedstawicielstwie wegetatywnym niż zwierzęcym.
Według Jaspera podstawowy ludzki rytm, tak zwany rytm alfa, wykazuje graniczną częstotliwość do 13 na sekundę, przeciętną zaś częstotliwość około 10 do 10,5 na sekundę, przy czym ta osobnicza przeciętna wykazuje dużą stałość, gdyż różnice z dnia na dzień nie przekraczają 10%. Graniczne wielkości amplitudy tych fal alfa wahają się od 5 do 100 mikrowoltów. Otrzymanie takiego normalnego ludzkiego encefalogramu związane jest z szeregiem warunków, a więc badany osobnik musi mieć powyżej 18 lat, nie może wykazywać choroby cielesnej lub psychicznej, musi pozostawać w stanie zupełnego czuwania, ale nie wykazywać żadnego napięcia ani cielesnego, ani umysłowego, musi mieć oczy zamknięte i w ogóle musi być możliwie wolny od podrażnień, mącących spokój. Każdy z wymienionych tu czynników wpływa na rytm wyładowań, które cechują się taką zmiennością 1 nieregularnością, jaką stwierdzamy znów tylko w. pobudliwości układu wegetatywnego, np. podczas badania chronaksji przedsionkowej, która pomimo podobnej zmienności występującej pod wpływem czynników, wykazuje jednak dość znaczną stałość osobniczą, począwszy — znów podobnie do normalnego encefalogramu — od wieku pokwitania poprzez cały okres dojrzałości.
Uwzględnianie wieku osobnika badanego jest konieczne dlatego, że u ludzkich niemowląt autorzy nie mogli wykryć w ogóle żadnych potencjałów mózgowych. Najmłodsze niemowlę, u którego Berger je wykazał, miało 35 dni, przy czym amplituda fal była niska, trwanie fal 160 sigm, a więc dłuższe niż normalnych fal alfa (90 do 120 sigm), następstwo bardziej nieregularne. Z wiekiem — od 1 miesiąca do 5 lat — wielkość potencjałów wzrasta, fale stają się krótsze i regularniejsze. Na ogół zbliżone wyniki otrzymał Lindsley, który stwierdził, że rytmiczne fale alfa, zawsze najlepiej zbierane w okolicy potylicznej, początkowo przypadkowe, rozwijają się u dziecka w wieku między 3 a 6 miesiącem, a więc w okresie, gdy dziecko zaczyna spostrzegać przedmioty, co zdaje się wskazywać na związek przyczynowy, przy czym wykazują częstotliwość 4 do 5 na sekundę, która potem stopniowo wzrasta i dochodzi u dziecka w wieku 8 lub 9 lat do wielkości stwierdzanej u dorosłego. Powolne rytmy i przypadkowe potencjały, podobne do stwierdzanych u dzieci, były spostrzegane także u dorosłych w warunkach patologicznych, albo pod wpływem farmakologicznych środków uspokajających, albo wreszcie podczas snu, w czasie którego niekiedy wszystkie potencjały zostają tak zahamowane, że z trudnością dają się wykryć i na otrzymywanym rejestrze zbliżają się do linii prostej.