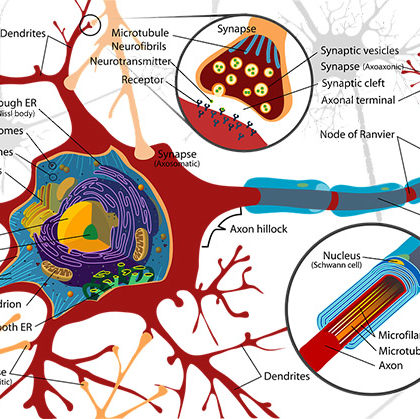
Berger obserwował w swych krzywych drugą serię potencjałów, które zwykle nakładają się na potencjały alfa albo trwają po zaniku rytmu alfa i które nazwał falami beta. Amplituda fal beta wynosi około czwartej części amplitudy fal alfa (zwykle poniżej 20 mikrowoltów), częstotliwość fal beta wynosi przeciętnie 25 na sekundę, waha się w granicach między 17 a 30 na sekundę . Ten rytm beta jest przeważającą cechą EEG u niektórych osobników, wykazujących bardzo małe fale alfa, które zdają się przeważać w przednim biegunie mózgu. Aby je można było wykryć poprzez czaszkę, muszą te fale być dostatecznie wysokie (u niektórych osobników fale te są poniżej dolnych granic niektórych aparatów rejestracyjnych).
Spór o to, czy fale beta są produkowane przez inne komórki nerwowe niż fale alfa, nie jest dotąd zakończony. W każdym razie z naszego punktu widzenia, który w przednim i tylnym biegunie półkul mózgowych upatruje dwa hierarchicznie różne narządy nerwowe, jest sprawą bardzo ciekawą, że wielu poważnych autorów widzi w tych dwóch biegunach kory wytwarzanie odrębnych potencjałów. Tak więc Adrian skłonny jest upatrywać w okolicy potylicznej jedyne źródło rytmu alfa. Adrian i Yamagiwa stwierdzili, że największy potencjał występuje w okolicy potylicznej, najmniejszy zaś potencjał w okolicy czołowej. Jasper, mówiąc o lokalizacji, twierdzi, że »maksymalną amplitudę i regularność spostrzega się na potylicy, niższa zaś amplituda i mniejsza ilość fal jest zazwyczaj regułą przy ich badaniu z przedniego bieguna głowy, chociaż istnieją niektóre wyjątki z tej reguły nawet u pozornie zdrowych osobników». Również Davis i Davis sądzą, że rytm alfa przejawia się przede wszystkim w okolicy potylicznej, rytmy zaś beta wiążą się z nerwową czynnością komórek przedniej części kory.
Najdalej pod względem lokalizacyjnym posuwa się Kornmüller, który sądzi, że rejestruje odrębne rytmy nawet z poszczególnych pól cytoarchitektonicznych. Krańcowo przeciwne zapatrywanie wypowiada Z. Drohocki, według którego »filtrowane» obrazy elektroencefalogramów przemawiają za tym, że w procesach korowych brak jest jakiejkolwiek ciągłości i że ich najwybitniejszą cechą jest właśnie nieregularność krzywych EEG. Jasper i Andrews potwierdzają spostrzeżenie Bergera, że rytm’ alfa o częstości 10 na sekundę może powstawać z innych okolic kory, nie tylko z potylicy, i sądzą, że właściwie należałoby mówić o politycznym, precentralnym, czołowym itd. rytmie alfa, bo chociaż zwykle jest duże podobieństwo częstotliwości i często jest dobra synchronizacja, to jednak w pewnych warunkach i u pewnych osobników regionalne rytmy alfa mogą się okazywać zupełnie niezależne.
Te w wysokim stopniu złożone i lokalizacyjnie dotąd jeszcze nie ustalone stosunki wikłają się jeszcze bardziej, jeżeli uwzględnimy opinię autorów, którzy twierdzą, że »różnice w budowie kory mózgowej zdają się mieć mniejsze znaczenie» niż niektóre inne czynniki, ponieważ osobniki, które zwykle nie wykazują żadnego rytmu alfa albo wykazują go bardzo mało, mogą dawać dobry regularny rytm alfa, jeżeli się przedsięweźmie wyjątkowe ostrożności, aby doprowadzić badanego do zupełnego zwolnienia napięcia nerwowego, albo jeśli badany znajduje się pod wpływem farmakologicznych środków uspokajających. Warunkowy, zależny od chwilowego stanu kory mózgowej charakter jej reakcji elektrycznej przejawia się najjaskrawiej w tych przypadkach, w których bodziec, zastosowany podczas snu, powoduje zjawienie się szeregu regularnych fal alfa, podczas gdy ten sam bodziec zastosowany w stanie czuwania może spowodować ich zahamowanie.
Jasper twierdzi, że fizjologiczna rola samoistnego, 10-cyklowego rytmu kory mózgowej, znajdowanego nad różnymi jej okolicami, bynajmniej nie jest dotąd wyjaśniona. Dużo przemawia jednak za tym, że rytm ten odgrywa w czynnościach korowych jakąś rolę podstawową. Wynika to zarówno z jego wszechobecności w różnych częściach kory mózgowej jak i z jego stosunku do impulsów odśrodkowych. Mianowicie Horsley i Schafer już w roku 1881 znaleźli, że kurcze mięśniowe, wywoływane przez podrażnienia korowe u psów, kotów, królików i małp, wykazywały rytm niezależny od częstotliwości po-drażnienia i wynoszący przeciętnie 10 na sekundę (w granicach między 8 a 13 na sekundę), a więc dający cyfry ściśle odpowiadające korowemu rytmowi alfa u człowieka. Zresztą Schafer wykazał znowuż te same cyfry (8 do 13 impulsów nerwowych na sekundę, przeciętnie 10) u człowieka w jego kurczach mięśniowych dowolnie wywoływanych. Przed kilku laty (1936) Jasper i Andrews rejestrowali równocześnie ruchy drżenia palca i potencjały akcji mięśniowej, a także z okolicy precentralnej u człowieka, i znaleźli w tych rejestrach drżenia 2 składniki: większy rytm (około 20 do 30 na sekundę), co ściśle odpowiada składnikom alfa i beta potencjałów mózgowych. Normalnie podkorowa kontrola rytmów kurczowych jest synchronizowana z kontrolą korową do mniej więcej tej samej częstotliwości, ale w pewnych warunkach (podrażnień kory lub jej uszkodzeń) mechanizmy podkorowe mogą górować.
Dziesięciocyklowy samoistny rytm korowy zdaje się być zatem rzeczywiście jakimś dynamizmem elementarnym, podstawowym, dla kory mózgowej powszechnym i udzielającym się nawet odśrodkowym i obwodowym narządom ruchu. Tym dziwniejszą wydaje się rzeczą, że fizjologiczna rola tego podstawowego korowego rytmu jest dotąd zupełnie niejasna. Jasper w końcowych wnioskach swej monografii podkreśla zorganizowaną synchroniczność tych spontanicznych rytmicznych wyładowań i sądzi, że »ta rytmiczna ciągłość aktywności korowej, ściśle związana z kompleksową integracją dośrodkowych impulsów sensorycznych, nie jest w swym istnieniu zależna od układu dośrodkowego, ale służy za dynamiczny ośrodkowy pośrednik pomiędzy bodźcem a odpowiedzią nań, a także służy do odpowiedzi zainicjowanych centralnie». W całej pracy niniejszej wychodziliśmy z założenia, że dynamicznym ośrodkowym pośrednikiem pomiędzy bodźcem a odpowiedzią jest układ wegetatywny na wszystkich piętrach osi mózgowo-rdzeniowej, począwszy od odruchów rdzeniowych, a kończąc na »odpowiedziach zainicjowanych centralnie» z pewnymi zastrzeżeniami, jak w reakcjach instynktownych, albo bez zastrzeżeń, jak w czynnościach zwanych »dowolnymi». Okazuje się zatem, że dalsze poparcie tego poglądu znajdujemy znów tutaj, w tej najnowszej gałęzi wiedzy neurofizjologicznej, mianowicie w elektroencefalografii.
W poprzednich rozdziałach tej pracy staraliśmy się wykazać, że wegetatywno-nerwowe pośrednictwo między bodźcem a odpowiedzią nań organizmu na poziomie podkorowym ma charakter potrzeb i dążności ustrojowych instynktowych, silnie zabarwionych uczuciowo, na poziomie zaś korowym należy przypuszczać, że aktywność korowego układu wegetatywnego jest fizjologicznym odpowiednikiem uczuciowości korowej ewolucyjnie wyższej. Gdyby przypuszczenie to było uzasadnione, musiałoby, oczywiście, znaleźć potwierdzenie w jakimś bliższym stosunku potencjałów korowych do życia uczuciowego. Stosunek ten, niestety, nie jest jeszcze bliżej wyjaśniony. Ale bodaj większość autorów, zajmujących się elektroencefalografią, zwraca uwagę na wybitny wpływ stanów uczuciowych, wywierany na przebieg wyładowań elektrycznych w korze.
Zdjęcie pochodzi z portalu wikimedia.org. Autorem zdjęcia jest Andrii Cherninskyi. Zdjęcie zostało wykorzystane na podstawie licencji CC BY-SA 4.0.