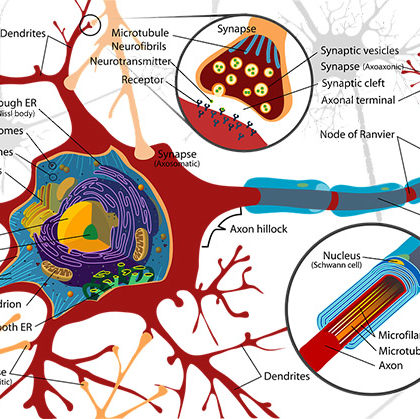
Berger sądzi, że fale alfa krótkie i o niskiej amplitudzie są »objawem podniecenia» czynności korowej, obecność zaś dużych i powolnych fal jest »objawem porażennym». Zahamowanie rytmu alfa, występujące zazwyczaj w warunkach normalnych pod wpływem działania jakiegoś bodźca, uważa Berger za prawdziwe zahamowanie aktywności większej części kory, wywołane jako reakcja na wzmożoną aktywność danego ogniska, które jest fizjologicznym odpowiednikiem »ogniska uwagi», jej »skupienia» na bodziec działający /. zewnątrz. Powtarzanie się podniety zwykle pomniejsza jej wpływ na rytm alfa albo go w ogóle usuwa, również sprawia to brak »zwracania uwagi» na podnietę.
Berger wnioskuje, że uwaga jest tu ważniejsza niż cechy samego bodźca jako, ta kiego. Rytm fal alfa jest według Bergera wyrazem aktywności psychofizycznej, odpowiednikiem stanu świadomości, stanu podniecenia bardzo bliskiego do procesu uwagi, którego pozytywny m wyrazem są fale alfa. Znikanie tych fal pod wpływem wysiłku uwagi sprzeciwia się, według Bergera, tylko pozornie powyższemu poglądowi, ponieważ podrażnieniu każdego zlokalizowanego ośrodka towarzyszy takie zahamowanie wszystkich innych ośrodków, którego pozornym następstwem jest osłabienie lub zanikanie fal alfa.
Zupełnie inaczej ujmują znaczenie fal alfa Adrian i Matthews, którzy widzą sprzeczność pomiędzy poglądem Bergera a faktami, zwłaszcza zaś faktem trwania, a nawet nasilania się fal alfa u ludzi, pozostających pod nasennym działaniem środków barbiturowych. Badacze angielscy podkreślają, że źródłem tworzenia się fal alfa jest okolica potyliczna, której czynność nić zanika nigdy zupełnie nawet w okresie »spoczynku», nawet przy zamkniętych oczach, gdy przyczyny aktywności elektrycznej działają w sposób jednostajny na elementy korowe, wskutek czego rytmiczne potencjały łatwo się synchronizują, a nakładając się dobrze jedne na drugie umożliwiają wykrycie potencjału globalnego. Natomiast synchronizacja staje się niemożliwa z chwilą, gdy wzrokowy ośrodek spostrzegania rozpoczyna swą czynność, ponieważ spostrzeganie kształtów wymaga uczynnienia bardzo wielkiej liczby elementów. Tak więc w ujęciu Adriana i Matthewsa zanikanie fal alfa jest wyrazem nie zahamowania, ale aktywnej fazy czynności kory.
Lemere znajduje mniej krzywych z »dobrymi» falami alfa u uczuciowo otępiałych schizofreników aniżeli u chorych maniakalno- depresyjnych i stąd wnioskuje, że główna przyczyna różnic w potencjałach wynika z różnic życia emocjonalnego. Jasper pisze, że »myśl o poziomach stanu podrażnieniowego kory albo aktywności korowej jest koncepcją raczej mglistą, ale okazała się pożyteczną hipotezą roboczą przy opisywaniu pewnych systematycznych zmian we wzorach potencjałów korowych, związanych ze stanami ustroju, zazwyczaj określanymi jako »podniecenie», »pobudzenie», »napięcie emocjonalne», »duża czujność», »silna koncentracja» itp. w przeciwstawieniu do zwolnienia napięcia, senności i snu. Większość warunków patologicznych, powodujących zaburzenie lub zahamowanie funkcji korowej, wiąże się z powolnymi falami, podczas gdy ostre stany podniecenia wiążą się z potencjałami o bardziej szybkiej i niskiej amplitudzie. Takie zmiany zachodzą u osobników normalnych w razie zmiany poziomu ich drażliwości ogólnej. Największa regularność i amplituda rytmu alfa u człowieka jest spostrzegana w optymalnych warunkach zwolnienia napięcia. Silne troski, podniecenia emocjonalne albo jakieś złe przeczucia często dają widoczne zahamowanie rytmu alfa, wyjąwszy rzadkie grupy fal w długim rejestrze. Ale i Jasper podkreśla, że podstawowy charakter tej ogólnej drażliwości nie jest jasny i zależy od wielkich indywidualnych różnic dążności do trwałego, regularnego rytmu alfa. Niektórzy ludzie wykazują wzmożone napięcie mięśniowe i podają, że są niespokojni lub zatroskani o coś, a jednak wykazują dobry rytm alfa, podczas gdy inni, którzy zdają się być zupełnie wolni od napięcia, mogą wykazywać bardzo mały rytm alfa. Jasper przypuszcza, że te różnice indywidualne mogą się sprowadzać do pewnej postaci przewlekłego podrażnienia korowego, w którym jednak odróżnienie stanu »napięcia» od stanu »zwolnienia» staje się już trudne. Na ogół jednak częstotliwość rytmu alfa zmniejsza się o 12 do 20% wraz ze zmianami stanu pobudzenia od »czujności» do »zwolnienia napięcia umysłowego» lub do »senności».
Adrian i Matthews, mówiąc o tym, że otwarcie oczu i inne nagłe bodźce wzrokowe są najbardziej niezawodnym środkiem zahamowania ludzkiego rytmu alfa, podkreślają, że ten rytm ulega szczególnie szybkiemu zahamowaniu, jeśli na obwodzie jednostajnie oświetlonego ekranu przesuwany jest jakiś przedmiot, który zwraca uwagę badanego, pomimo polecenia, aby się wpatrywał w środkowy punkt ekranu. Jeżeli przesuwany przedmiot uwagi nie zwraca, rytm alfa nie ulega żadnej zmianie. Ci autorzy podkreślają w ogóle wielką rolę »zaskoczenia», działania nieoczekiwanego, np. pytania postawione w sposób niespodziewany powodują zanikanie fal alfa, natomiast zwykła rozmowa, nie wymagająca żadnego poważnego wysiłku uwagi, albo jakaś zwykła, przeciętna praca mięśniowa, nie wywołują wyraźniejszego wpływu na rytm alfa. W dalszych doświadczeniach Adrian i Yamagiwa wychodzili, jak to już mówiliśmy, z założenia, że u człowieka normalnego źródłem fal alfa jest okolica potyliczna i poszukiwali odpowiedzi na pytanie, czy określony wysiłek myślowy nie może zmienić topografii źródła rytmu alfa. Badany otrzymał polecenie napisania liter alfabetu mając zamknięte oczy, a więc w warunkach niezwykłych, które istotnie spowodowały zanik fal alfa przez czas tego pisania. Podczas trzeciej takiej samej próby fale alfa już nie uległy zmianie, tylko spostrzeżono przemieszczenie ogniska w kierunku bieguna potylicznego. U dwóch innych badanych, którzy zmuszeni zostali do ciężkiego wysiłku umysłowego, ognisko przemieściło się w kierunku linii środkowej; jakość wysiłku umysłowego okazała się bez znaczenia.
Durupt i Fessard, przeciwnie, bardzo silnie podkreślają znaczenie uwagi w tych przypadkach i twierdzą, że bez uwagi wzrokowej, np. w razie jej ześrodkowania na podniety słuchowe, drażnienie siatkówki przez świetlne wzory nie miało żadnego wpływu na rytm alfa. Inni autorzy (Loomid, Harvey i Hobart 1936) znaleźli, że »w hipnozie fale alfa trwały w dalszym ciągu, jeżeli badany miał oczy otwarte, ale mówił, że nie widzi, natomiast ulegały zahamowaniu za każdym razem, gdy mówił, że widzi». Ci sami i inni autorzy podkreślają, że duży wysiłek umysłowy, np. rozwiązywanie jakiegoś zagadnienia, hamuje rytm alfa w tych przypadkach, w których się wiąże z elementem »napięcia emocjonalnego», obawą błędnego rozwiązania itp. Tę zależność rytmu alfa od czynników wzruszeniowych, związanych z aktem uważania, tłumaczą niektórzy autorzy (Berger, Jasper i Andrews) tym, że przypuszczalnie rytm ten jest »dyktowany» korze z okolicy podkorowej.
Przewagę podniet wzrokowych u człowieka pod względem hamowania fal alfa trzeba uzależniać, zdaniem Jaspera, raczej od znaczenia wzroku na wyższych poziomach encefalizacji czynności aniżeli od swoistego stosunku rytmu alfa do mechanizmu wzrokowego jako takiego. U szczura najsilniej blokuje potencjały korowe dźwięk, naśladujący pisk szczura; u kota bodźce dotykowe i słuchowe silniej wpływają na zahamowanie powolnych rytmów aniżeli bodźce wzrokowe.
A wreszcie Jasper z Andrewsem, a także z Rheinbergerem stwierdzili eksperymentalnie istnienie dwóch kategorii reakcji: jedną kategorię kompleksowych, »masowych» odpowiedzi całej kory na bodźce, które powodują irradiację aktywacji na całą korę i wywołują uogólnioną działalność kory i drugą kategorię reakcyj dobrze zlokalizowanych, np. ograniczających się do jakiegoś pola cytoarchitektonicznego i występujących pod wpływem bardziej prostych bodźców, zwłaszcza wtedy, jeśli uogólniona aktywacja została stłumiona przez lekkie znieczulenie dialowe. W tym podziale reakcji na »zlokalizowane» i »masowe całej kory» widzimy znów nie tylko tę samą dwoistość czynnościową układu wegetatywnego, którą omawialiśmy w rozdziałach 0 jego »jedności» i jego zróżnicowaniu, ale także tę dwoistość, którą widzieliśmy tak w dynamizmach podrażnień ograniczonych pozornie do luku odruchowego jak i w dynamizmach »masowych», swoistych i odrębnych dla każdego piętra osi mózgowo-rdzeniowej. Naturalną, biologiczną odpowiedzią są tylko te reakcje »masowe», które odtwarzają poszczególne stadia rozwoju gatunkowego i osobniczego; odpowiedzi odruchowe, izolowane są tylko sztucznym produktem eksperymentu (Sherrington, Goldstein i inni). Również Adrian mówi — jak przed chwilą widzieliśmy — o »potencjale globalnym», powstającym przez synchronizację i nakładanie się rytmicznych potencjałów jednych na drugie, mianowicie w tych przypadkach, w których »przyczyny aktywności elektrycznej działają w sposób jednostajny na elementy korowe». Ale na ogół w znaczeniu ewolucyjnym różnych potencjałów korowych wcale nie możemy się jeszcze orientować.