Przytoczone w tym rozdziale wyniki badań doświadczalnych są bardzo niepełne, ale w ogólnych zarysach dają już jednak pewne wyobrażenie o tym, że neurodynamizmy obwodowe i ośrodkowe, nawet najwyższe, mianowicie korowe i korowo-psychiczne, jak praca umysłowa, będąca najwyższym kierownictwem ustroju, odbywają się z najmniejszym wyładowaniem energii, pozostającym w rażącym niestosunku do biologicznego znaczenia procesów psychicznych. Zresztą nie może być inaczej, jeżeli słuszny jest nasz pogląd, że wszystkie procesy korowo-psychiczne są grą swoistych nastawień, początkowo należących do odruchowości warunkowej, a powstających przez tworzenie się nadbudowy mnemiczno-energetycznej, czyli przez przekształcanie się chronaksji konstytucyjnej w chronaksję subordynacyjną. Pogląd ten zdają się potwierdzać po pierwsze doświadczalne badania Monniera i Jaspera, z których wynika, że przekształcanie się chronaksji konstytucyjnej w subordynacyjną, dające nastawienie wtórne, odbywa się pod wpływem najmniejszego zwiększenia dodatniego ładunku elektrycznego (komunikat Lapicque’a na XV Międzynarodowym Zjeździe Fizjologów w Moskwie); po drugie francuscy autorzy wykazali, że podczas przebiegu odruchu warunkowego zmienia się jednakowo chronaksja nerwów obwodowych i komórek kory mózgowej, mianowicie początkowo wysokie chronaksję obniżają się podczas wykonywania ruchu i znowu wzrastają po skończonym odruchu. Rosyjscy autorzy, Levitina i Fasler stwierdzili, że u nietrenowanych potencjał skórny spada podczas pracy tylko miernie, w porównaniu do chronaksji ruchowej, ale podnosi się powoli. U ludzi trenowanych, przeciwnie, spada silnie, ale za to szybko podnosi się z powrotem. U chorych, którzy ulegli wycięciu nerwu współczulnego, autorzy ci spostrzegali po stronie pozbawionej nerwu współczulnego znaczne obniżenie chronaksji, podczas gdy na drugim ramieniu chronaksja zwiększa się pod wpływem pracy. Na zmiany chronaksji ruchowej wpływa zatem układ nerwowy wegetatywny.
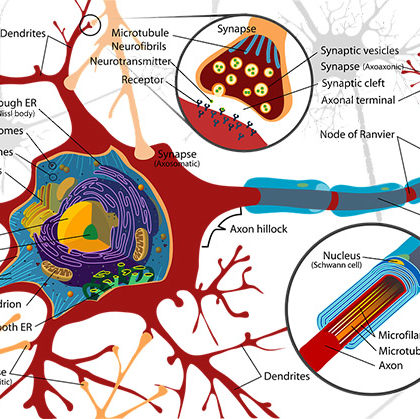
Autorzy, zajmujący się badaniem elektrycznej aktywności kory mózgowej, interesowali się przede wszystkim lokalizacją różnych potencjałów w różnych płatach lub nawet polach cytoarchitektonicznych. W dostępnej mi literaturze nie znalazłem innych prób lokalizacyjnych, możliwych zresztą tylko w znaczeniu hipotetycznym. Otóż można przypuszczać, że w korze mózgowej, oprócz dróg tzw. »kojarzeniowych» i spoidłowych, wiążących obie półkule ze sobą, istnieją jeszcze przedłużenia wszelkiego rodzaju dróg dośrodkowych, a więc epikrytycznych, proprioceptywnych i wegetatywnych- (afektywnych). Tym korowym przedstawicielstwom wszelkich układów dośrodkowych musimy przypisywać wbrew dawniejszym poglądom — rolę nie tylko przewodzącą i kojarzącą podrażnienia w poszczególnych neuronach, ale rolę anatomicznego podłoża, które przechowuje — dzięki engrafii — wszelkie doświadczenia życiowe i które je opracowuje, a więc anatomicznego podłoża pamięci osobniczej i wszelkich procesów psychicznych.
Pojęcia komórek nerwowych »pamięciowych» i »psychicznych» zostały dawno już gruntownie pogrzebane i ośmieszone, — nie wrócą nigdy. Ale trzeba powiedzieć wyraźnie, że reakcja przeciwko tym naiwnym anatomicznym lokalizacjom nie-istniejących elementów psychicznych wpada często w tak daleko idący przeciwległy kraniec, nie uznający zasadniczo w ogóle żadnej lokalizacji czynności psychicznych, że ta druga krańcowość z punktu widzenia psychofizycznej jedności organizmu i zwłaszcza z punktu widzenia psychofizjologii musi być uznana za równie szkodliwą i śmieszną jak pierwsza krańcowość. Nie jest żadną hipotezą, ale dzisiaj jest już stwierdzonym faktem biologicznym, że dynamizmy instynktowe są dynami/ mami podkorowymi i że płaszcz mózgowy jest na rządem pamięci osobniczej i procesów psychicznych. Tylko z tych faktów wychodząc, a nie z bezpłodnego przeczenia, wysiłki umysłu ludzkiego doprowadzą kiedyś do stworzenia w przyszłości psychofizjologii.
Dzisiaj w wielu, a właściwie w większości zagadnień psychofizjologicznych musimy poprzestać na przypuszczeniach; ale nawet przypuszczenia są korzystniejsze od czczego zaprzeczania, jeżeli opierają się na takich faktach i doprowadzają do takich poglądów, które są chociażby o jeden mały krok bliższe rzeczywistości niż dawniejsze doktryny. Do takich owocnych hipotez zaliczamy zainicjowaną przez Se mona, Monakowa i Mourgue’a chronogenną lokalizację zasobów mnemicznych, pomimo tego, że fizjologicznie nie możemy jej sobie dotąd nawet wyobrazić. Drugą pożyteczną hipotezą, którą my proponujemy i która zdaje się być daleko bliższa dzisiejszej rzeczywistości niż poglądy klasyczne, jest ujęcie w sposób masowy globalny swoistych czynności każdego poszczególnego piętra układu nerwowego jako jedności czynnościowych, stwierdzanych doświadczalnie na poziomie segmentów rdzenia kręgowego i na poziomie kory pozaczołowej. A wreszcie do takich owocnych hipotez zaliczamy uogólnienie W. R. Hessa i Orbeliego o kierowaniu całego układu nerwowego zwierzęcego przez układ wegetatywny, uogólnienie, które w naszym ujęciu znajduje zastosowanie, wprawdzie hipotetyczne, ale konsekwentne i — jak się zdaje — uzasadnione w stosunku do wyższych pięter układu nerwowego, do ewolucji jego czynności i czynności psychicznych, których motorem dynamicznym — na wszystkich poziomach ewolucji — jest życie uczuciowe, które w fizjologii musi mieć swój odpowiednik neurodynamiczny. Odpowiednik ten na poziomie podkorowym poznaliśmy pod postacią czynności okresowej »ośrodków potrzeb ustrojowych» a na poziomie korowym — skłonni jesteśmy hipotetycznie upatrywać go w czynnościach jej przedstawicielstwa wegetatywnego, jako odpowiednikach wyższych stopni rozwojowych życia uczuciowego. Otóż w tym miejscu musimy raz jeszcze przypomnieć bodaj że powszechnie uznawane twierdzenie fizjologów, które przeciwstawia stan spoczynku, cechujący układ nerwowy zwierzęcy1, dopóki nań nie zacznie działać jakaś podnieta, — stanowi nieustannej aktywności układu nerwowego wegetatywnego, który stanu spoczynku w ogóle nie zna, który go nawet znać nie może, ponieważ z natury swej jest związany z układem gruczołów dokrewnych, wywierającym nań wpływ również ciągły, jak ciągłą jest kontrola układu nerwowego wegetatywnego nad czynnością układu gruczołów dokrewnych i wielkich funkcji ustrojowych; ustanie aktywności układu wegetatywnego byłoby zatem ustaniem przemiany materii w ustroju, ustaniem jego życia. Trzeba przyznać, że w tych warunkach trudno sobie wyobrazić, aby samorzutna aktywność potencjałów korowych, o których była mowa w tym rozdziale i które niezupełnie zanikają nawet podczas snu, mogła być działalnością korowego przedstawicielstwa układu zwierzęcego, skoro ta działalność nigdy nie jest samorzutna, rytmiczna i ciągła. Oczywiście, czynność układu wegetatywnego może być również odruchowa na wszystkich piętrach układu nerwowego, samorzutna zaś — a właściwie rzekomo samorzutna aktywnością własną może się stawać tylko na najwyższych poziomach rozwoju czynności psychicznych i korowo-mnemicznych.
To, co Adrian nazywa »potencjałem globalnym», Jasper »potencjałem masowym», zdaje się być zatem korową postacią piętrowych jedności czynnościowych, których mniej złożone postacie poznaliśmy na poziomie podkorowym (jedności czynnościowe, kierowane np. przez energetycznie naładowane ośrodki potrzeb fizjologicznych) i na poziomie rdzeniowym (segmentarne jedności czynnościowe Dusser de Barenne’a, Heada i innych). Na wszystkich tych poziomach anatomicznym elementem jednoczącym jest układ wegetatywny, a twierdzenie Adriana i innych autorów, że potencjał globalny przedniego bieguna kory mózgowej różni się od potencjału jej bieguna tylnego, zdaje się być jednym więcej dowodem tego, że w korze mózgowej należy upatrywać nie jeden, ale dwa narządy o odmiennych czynnościach globalnych.
Wreszcie przypomnimy tu raz jeszcze, że u noworodków nie wykazano wcale tych potencjałów, że zaczynają się one rozwijać w wieku między 3 a 6 miesiącem i wzrastają według jednych do 5 roku życia, a według innych (Lindsley) — do 8—9 roku życia, w którym dochodzą do wielkości stwierdzanej u dorosłego. Jeżeli równocześnie przypomnimy sobie w tym miejscu stwierdzone przez Skrzypińską zmniejszanie się pobudliwości konstytucyjnej układu przedsionkowego w cyklu życiowym człowieka, to wzrastanie potencjałów korowych u dziecka, świadczące o wzmaganiu się pobudliwości nabytej subordynacyjnej kory mózgowej, zdaje się wskazywać wyraźnie na mnemiczny charakter tej zmiany.
Zdjęcie pochodzi z portalu wikipedia.org. Źródło zdjęcia:
BMC Infectious Diseases 2006. Zdjęcie zostało wykorzystane na podstawie licencji CC BY 2.0.